just read an article on the web. a good read, really quite a good read, especially during such suffocating times.
剛在網上讀到一篇文章。好讀,真的是很好的好讀,尤其是在現在這些令人窒息的時間之中。
these days, it is really rare to find an informative and inspiring journal based on thorough and in-depth fact-based researches and studies.
今時今日,很難得會找到一份建基於詳盡,深入及以事實為基礎,而又具備資訊性和啟發性的調查研究。
allow me to list some extracts from it first, then you’ll know when i am talking about. before sharing with you the complete text.
容讓我先列出一些摘錄,那你就會明白我在說甚麼。就在我跟你分享那全文之前。
@@@@@@@@@@@@@
…It grows and may die. It reacts to varying environmental conditions and other organisms, producing different growth forms or structures, depending on circumstances…
……它會生長並可能死亡。 它會對不同的環境條件和其他生物做出反應,並根據情況產生不同的生長形式或結構……
…while the hyphae show a very marked outward growth, there are also cross-connections between the outward growing branches…
……雖然菌絲顯示出非常明顯的外向生長,但向外生長的分支之間也存在交叉連結……
…the mycelium actually grows as an expanding ring, not as an expanding disk…
……菌絲體的生長實際上是一個擴展環而不是一個擴展盤……
…the mycelium produced from a single spore will not produce fruiting bodies. It is necessary for two mycelia to mate and the resulting, expanding mycelium can then produce the fairy-ring mushrooms…
……單個孢子產生的菌絲體不會產生果體。必須有兩個菌絲體交配後和由此產生的,擴展的菌絲體才能生產出精靈環蘑菇……
…the ring-like underground mycelium has little or no visible effect and only makes its presence known when it produces the visible fruiting bodies…
……環狀地下菌絲體幾乎或完全沒有可見的蹤跡,只有當它生產出可見的果體時才會知道它的存在……
…the underground mycelium secretes chemicals that break down organic matter in the soil to release nutrients for the fungus to use. For a short time the grass just beyond the outer edge of the mycelium can also make use of these nutrients…A little way behind the outer edge, the mycelium is much denser, actively feeding on the liberated nutrients and leaving little for the grass…reducing the water available to the grass and so putting further stress on the grass. Further inwards, there are the remnants of the cannibalized mycelium. This is broken down by various micro-organisms and the resulting nutrients are released into the soil. This increase in nutrients behind the mycelial ring explains the enhanced grass growth immediately behind the ring of brownish grass. Therefore the brownish grass recovers as the ring expands…
地下菌絲體生長的前沿會分泌化學物質分解土壤中的有機物質,釋放出可供真菌使用的養分。在一段短時間內,菌絲體外圍以外的草也可以利用這些養分……在外圍後面一點的位置,菌絲體密度要大得多,積極地吸收已釋放的養分,留下給草可吸收的會很少……再往裡面,殘餘了被拆解的菌絲體。這會被各種微生物分解,而所產生的養分會被釋放到土壤中。菌絲環後面養分的這增加解釋了緊接在啡色草環後面的草的生長有所增強。因此,隨著環的擴大,啡色的草會恢復……
…A sclerotium is a resting structure that allows a fungus to sit out the hard times, analogous to (but clearly different from) a plant bulb or corm. The outer skin protects the internal hyphal mass from drying out. When conditions improve, the sclerotium may produce a new mycelium or fruiting bodies…
……菌核是一種休止結構,可以讓真菌度過艱難時期,類似於(但明顯不同於)植物的鱗莖或球莖。外皮保護內部菌絲團免於乾涸。當環境改善時,菌核可能會產生新的菌絲體或果體……
…In most cases, hyphae grow out of the sheath into the delimitated chunk of substrate and slowly transform the latter into an intimate mixture of fungal biomass and residual substrate…
……在大多數情況下,菌絲從覆套中向外生長到認定的基底堆,再緩慢地把它們轉化為真菌生長所需的生物物質群和殘餘基底的緊密混合物……
…once a mycelium has spread to a considerable size it becomes more likely that it will suffer disruptions and fragmentation…leaving isolated fragments where once there was a continuous mycelium…
……一旦菌絲體擴散到相當大的規模,它就更有可能遭受破壞和碎片化……在曾經一體連的結菌絲體的地方留下孤立的碎片……
…The five parts of the original mycelium can continue growing independently…it is possible that at some future stage two of these currently disconnected mycelia will come into contact. Then, since genetically identical mycelia can fuse, the two mycelia can re-join…
……原本菌絲體分裂出的五個部分可以繼續獨立生長……然後,由於基因相同的菌絲體可以融合,兩個菌絲體可以重新結合……
…the mycelium as a heterogenous army of hyphal troops, variously equipped for different roles and in varying degrees of communication with one another. Without a commander, other than the dictates of their environmental circumstances, these troops organise themselves into a beautifully open-ended or indeterminate dynamic structure that can continually respond to changing demands…
……菌絲體是一支由菌絲組成的異質軍隊,裝備各異,用於不同的角色,彼此之間的交流程度也有所不同。沒有指揮官,除了它們身處的環境條件的規限,這些部隊將自己組織成一個美麗的開放式或不確定的動態結構,可以不斷回應不斷變化的需求……
interesting? truly, let us dig into it…now
有趣嗎?認真地,讓我們挖掘下去吧……就現在
While we may admire a mushroom growing out from the soil or a bracket fungus growing out from a log, it is the out-of-sight (and often forgotten) mycelium that is the essential part of the organism. The mycelium is there throughout the year, in the soil or in the log, and is not a static object. It grows and may die. It reacts to varying environmental conditions and other organisms, producing different growth forms or structures, depending on circumstances.
雖然我們可能會欣賞從土壤中長出的蘑菇或從原木中長出的支架真菌,但看不見(並且經常被遺忘)的菌絲體才是有機體的重要組成部分。 菌絲體一年四季都存在,在土壤中或在原木中,並且不是靜止的物體。 它會生長並可能死亡。 它會對不同的環境條件和其他生物做出反應,並根據情況產生不同的生長形式或結構。
This section will discuss some of the features of the mycelium.
本節將討論菌絲體的一些特徵。
Hyphal growth
菌絲生長
Consider a spore from a simple mushroom, out in a field. The spore germinates and produces a short, initial hypha (called a germ tube). The germ tube grows and branches, each of those early branches grows and branches in turn and the process continues. The following diagram shows this process.
考慮來自野外的簡單蘑菇的孢子。 孢子萌發並產生短的初始菌絲(稱為胚芽管)。 胚芽管生長和分支,每個早期分支依次生長和分支,而這過程繼續發生。 下圖顯示了這個過程。
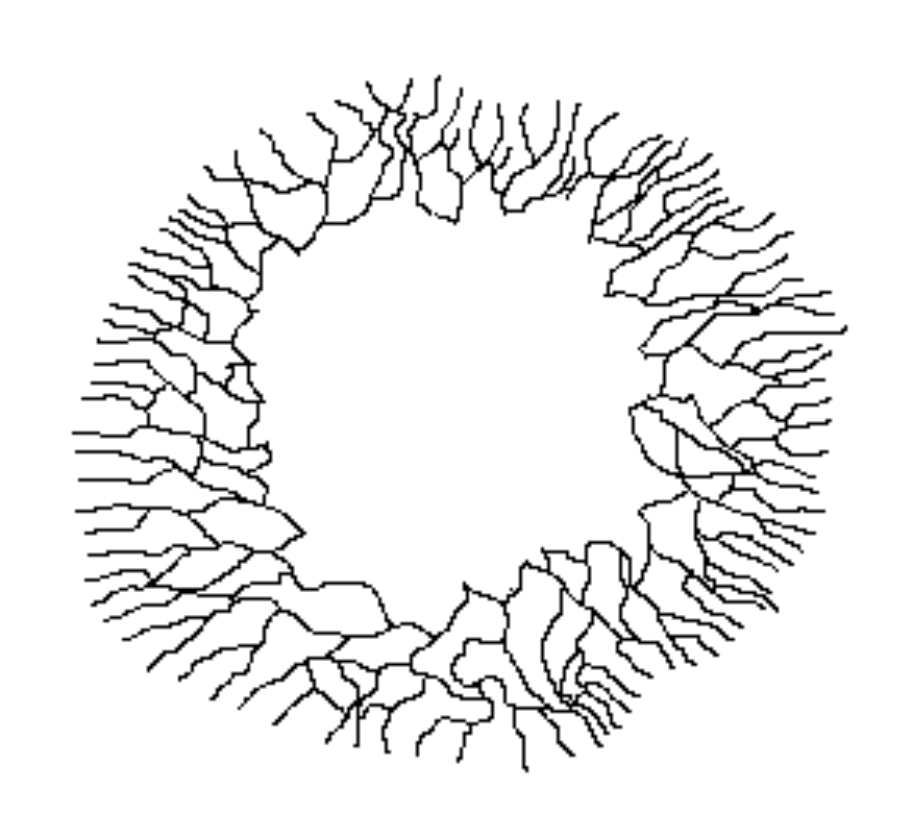
The leftmost figure shows a spore (the black dot) with the short germ tube growing out from it. The next figure shows the scene a little later, with several branchings having occurred by now. The other two figures show later stages in the expansion of the mycelium. Notice how, by repeated branching, the mycelium eventually assumes a circular form as shown in the rightmost figure. That figure also shows that while the hyphae show a very marked outward growth, there are also cross-connections between the outward growing branches. The cross-connections between the radiating hyphae make it easy to move nutrients quickly around the growing mycelium, taking them to wherever they are most needed.
最左邊的圖顯示了一個孢子(黑點),短的胚芽管從中長出。 下一個圖顯示了稍後的情況,此時已有幾個分支出現。 另外兩個圖顯示了菌絲體擴張的較後階段。 請注意,經過重複的分支,菌絲體最終呈現出如最右圖所示的圓形形態。 該圖還表明,雖然菌絲顯示出非常明顯的外向生長,但向外生長的分支之間也存在交叉連結。 輻射菌絲之間的交叉連結使營養物質可以輕易地在生長中的菌絲體各處快速移動,將它們帶到最需要的地方。
This website deals with the macrofungi and so microfungi such as the various moulds aren’t included. However, it’s worth noting that the common household moulds on bread and fruit provide excellent examples of the radial growth that has just been explained. These mould colonies are also composed of hyphae and are far more easily seen than the out-of-sight mycelium of a mushroom. Radial growth is a feature common to all fungi (macro and micro) that grow as hyphae. The yeasts are examples of non-hyphal microfungi, being single-celled and budding off daughter cells.
本網站探討的是大型真菌,因此並不包括各種黴菌等微型真菌。 然而,值得注意的是,家居常見的麵包和水果上的黴菌提供了剛剛說明的徑向生長的極好例子。 這些黴菌菌落也由菌絲組成,比看不見的蘑菇菌絲體更容易看到。 徑向生長是所有菌絲生長的真菌(宏觀和微觀)的共同特徵。 酵母菌是非菌絲微真菌的例子,它們是單細胞並且是從子細胞中萌芽的。
In many hyphal species, the individual hyphae in a mycelium are no more than a hundredth of a millimetre in diameter. A hypha grows through the addition of new material to the hyphal wall at or near the tip of the hypha and this reference button goes into technical detail about the mechanics of tip growth. A hypha grows by increasing in length. There is no increase in diameter (as in a plant root for example) but, as shown in the diagram above, hyphae branch. Branching normally occurs some distance behind the tips, so that the margin of a growing mycelium is typically composed of unbranched leaders, growing into fresh territory.
在許多菌絲物種中,菌絲體中的單一菌絲直徑不超過百分之一毫米。菌絲通過在菌絲尖端或附近的菌絲壁添加新材料而生長,此參考按鈕連結至有關尖端生長機制的技術細節。菌絲隨著增加長度而生長。而直徑沒有增加(好像植物的根一樣),但是,如上圖所示的菌絲分支。分支通常發生在尖端后面的一定距離,因此正在生長的菌絲體的邊緣通常由未分枝的前導組成,生長到新的區域。
The hyphae are not just uniformly featureless threads, the same in all species. For example, the hyphae may be smooth or encrusted, colourless or coloured and the hyphal walls may be two-layered to multi-layered. The hyphae of the macrofungi (and numerous microfungi) have cross-walls (or septa), which give structural support – and there are variations in septal structure. There are various chemical components to the hyphal walls, with chitin a significant component in the macrofungi. Insect exo-skeletons are also composed of chitin. While the prime purpose of a hyphal wall is to keep the inside in and the outside out, that need not be its only role. It can also be used as a place to store food reserves. For example, when it comes to producing fruiting bodies, the species Schizophyllum commune breaks down parts of the hyphal walls in the mycelium and uses the released nutrients in fruit body production.
菌絲不僅是均勻無特徵的線狀體,在所有物種中都是一樣的。例如,菌絲可以是光滑的或結殼的、無色或有色的,菌絲壁可以是兩層到多層。大型真菌(和許多微型真菌)的菌絲具有交叉壁(或隔膜),可提供結構性支撐——並且隔膜結構亦有變化。菌絲壁有多種化學成分,其中甲殼素是大型真菌的重要成分。昆蟲外骨骼也由甲殼素組成。 雖然菌絲壁的主要目的是在外部環境中保護內部,但這不一定是它的唯一作用。它也可以用作儲存食物儲備的地方。例如,在生產果體時,末麼茸會分解菌絲體中的部分菌絲壁,並把所釋放的養分用在果體的生產中。
In order to feed, the mycelium releases enzymes into the surrounding environment and these enzymes break down complex organic polymers into simpler compounds (generally various sorts of sugars) which are then absorbed through the hyphal walls. All this also happens near the growing hyphal apices, generally just behind the region where fresh material is being added to the hyphal wall. While this discussion uses a common mushroom as a simple example, it’s important to note that this method of feeding – enzyme release into the surrounding environment and absorption of the resulting products through the cell walls – is a fundamental characteristic of all fungi, whether macrofungi or microfungi.
為了取得食物,菌絲體向周圍環境釋放酵素,這些酵素將複雜的有機聚合物分解成更簡單的化合物(通常是各種醣類),然後透過菌絲壁吸收。所有這些也發生在生長中的菌絲尖附近,通常就在新鮮物質被添加到菌絲壁的區域後面。雖然這討論使用一些常見的蘑菇作為一個簡單的例子,但重要的是要注意這種攝取方法——將酵素釋放到周圍環境中並透過細胞壁吸收所產生的物質——是所有真菌的基本特徵,無論是大型真菌還是微型真菌。
Hyphal growth need not be random, since fungi respond to chemical signals. For example the hyphae of various parasitic or mycorrhizal species respond to chemicals that leak out from the tips of plant roots.
菌絲生長不必然是隨機的,因為真菌會對化學信號做出反應。例如,各種寄生或菌根物種的菌絲會對從植物根尖滲出的化學物質作出反應。
The combination of generally radial growth, with branching hyphae growing out from behind the leading hyphae, means that a mycelium can explore and exploit a large area. As the mycelium exhausts the food sources in one area and expands outward in a circular fashion, there’s no benefit in maintaining the inner mycelium. So the fungus cannibalizes the inner mycelium, extracting whatever nutrients it can from there and moves them to the outer, growing regions. Whatever cannot be recycled is shut off from the growing region and allowed to decay, so that the mycelium actually grows as an expanding ring, not as an expanding disk, and the accompanying diagram shows this.
結合了一般呈放射狀的生長、分支菌絲從前導菌絲後面長出,意味著菌絲體可以探索和開發一個大面積。當菌絲體耗盡了一個區域的食物來源並以圓環方式向外擴展,維持內部菌絲體沒有任何好處。因此,真菌會拆拼內部菌絲體,從那裡提取所有可獲取的養分,並把它們轉移到外部,生長中的區域。任何不能回收的東西會被從生長區域分離並由得其腐化,因此菌絲體的生長實際上是一個擴展環而不是一個擴展盤,附圖顯示了這一點。
If you’ve seen mushrooms growing in fairy rings in a field, this ring-like mycelial growth immediately suggests the cause. That is in fact the case, but one important point is that, generally speaking, the mycelium produced from a single spore will not produce fruiting bodies. It is necessary for two mycelia to mate and the resulting, expanding mycelium can then produce the fairy-ring mushrooms.
如果您曾在田野中看到過蘑菇在精靈環中生長,這種環狀菌絲生長可立即說明其原因。事實確實如此,但重要的一點是,一般來說,單個孢子產生的菌絲體不會產生果體。必須有兩個菌絲體交配後和由此產生的,擴展的菌絲體才能生產出精靈環蘑菇。
The earlier illustration of radiating mycelial growth showed a somewhat diffuse growth pattern, with individual hyphae growing out in all directions. If there is a fairly uniform distribution of nutrients in a benign environment, such radial growth is very efficient since it allows the fungus to make the most of those nutrients. However, nutrients are not always uniformly distributed and the surrounding environment is often hostile. These factors are major influences on the pattern of mycelial growth. If there’s a significantly higher concentration of nutrients in one area, then the mycelium would grow preferentially into that area, with less growth (or perhaps even none) into other areas – especially areas with a hostile environment.
之前的輻射性菌絲體生長圖顯示出某種程度的擴散生長模式,單個菌絲向各個不同方向生長。如果在養分分佈相當均勻的良性環境中,那麼這種徑向生長非常有效,因為它可以讓真菌充分利用這些養分。然而,養分並不總是均勻分佈,而且周圍的環境往往很不友善。這些因素是影響菌絲生長模式的主要因素。如果某一區域的養分濃度明顯較高,則菌絲體生長的會優先選擇進入該區域,而會較少生長(甚至可能生長)到其他區域——尤其是環境惡劣的區域。
In an area rich in nutrients, the mycelium will branch often and grow slowly, so maximising the amount of nutrients it can extract. By contrast, in an area low in nutrients the hyphae grow more rapidly and with little branching. Moreover, in many cases the bulk of the mycelium doesn’t grow as individual hyphae – but as bundles of hyphae. Some bundles are simply fairly loose and unstructured aggregations of hyphae, with no differentiation in the functions of the component hyphae. The opposite also occurs commonly, with a definite structure to the bundle and marked differentiation in the nature of the hyphae making up the bundle.
在養分豐富的區域,菌絲體會經常分支並緩慢生長,因此可以最大限度地提取養分。相比之下,在養分含量低的區域,菌絲生長得更快,而且很少分支。此外,在大多數的情況下,大部分菌絲體不會以單個菌絲的形式生長——而是以菌絲束的形式生長。一些束只是相當鬆散和無結構的菌絲聚合,組成中的菌絲的功能亦沒有區別。相反的情況也很常見,具有明確的束結構和組成束的菌絲的性質希顯著的差異。
Rhizomorphs 根狀莖
These hyphal bundles are referred to by various terms (mycelial cords, mycelial strands or rhizomorphs) and you will see some variation in the use of these terms. Some people use the term rhizomorph for anything but the loosest of bundles whereas others will use it only for highly structured bundles. I’ll use the word “rhizomorph” here as a general term for all such bundles. The reference given in the button contains a discussion of some of the terms that have been used for such hyphal bundles.
這些菌絲束有各種不同的術語(菌絲索、菌絲束或根狀體),您會看到這些術語的使用有所不同。有些人將術語根狀莖用於除最鬆散的束以外的任何型態,而其他人則只將其用於高度結構化的束。我將在此處使用「根狀莖」一詞作為所有此類束的通用術語。按鈕中連結的參考包含了對用於此類菌絲束的一些術語的討論。
The photo shows some white rhizomorphs growing through an area of wood chip mulch. You’ll often come across such rhizomorphs when raking aside dead wood or old mulch, while doing some gardening. There is considerable variation in both the structure and growth of rhizomorphs. In some species the rhizomorphs grow in a very regimented way, the tips extending as a whole, in a manner similar to the growth of root tips in plants. In other cases one or a few founder hyphae grow out and other hyphae then follow the same route as the founder hyphae and twine around them. In this way a bundle of hyphae builds up. In many species the outer regions of the mycelium grow in a diffuse, fanned-out fashion with rhizomorphs developing a little behind those expanding, diffuse extremities.
這照片 顯示了一些白色的根狀莖生長在一片木屑覆蓋的區域中。在進行園藝工作時,您經常會在耙掉枯木或長久覆蓋物時遇到這種根狀莖。根狀莖的結構和生長會存在相當大的差異。在某些物種中,根狀莖以非常有規律的方式生長,尖端整體延伸,其方式類似於植物根尖的生長。在其他情況下,一個或幾個原初菌絲向外生長,然後其他菌絲沿著與原初菌絲相同的路線並纏繞在它們周圍。這樣就形成了一束菌絲。在許多物種中,菌絲體的外部區域以散開、扇形的方式生長,而根狀莖在那些擴張、分散的端點的後面發育。
The following two diagrams will give you some idea of the features found in many of the highly structured rhizomorphs, with well regimented tip growth. But remember that there is variation in rhizomorph structure, so these are generalized diagrams, rather than faithful depictions of the rhizomorphs of any particular species.
下面的兩張圖可讓你了解到在許多高度結構化的根狀莖中可找到的特徵,這些根狀莖具有十分規律性的尖端生長。但請記住,根狀莖結構存在差異,因此這些是概觀圖像,而不是對任何特定物種的根狀莖的忠實描述。
The first diagram represents a cross-section, taken along a rhizomorph. The growing apex is on the right, where you can see the individual hyphae, intertwined but generally parallel and, in this area, the hyphae show no differentiation in function. A little way further back there is a marked difference in structure across the rhizomorph. Moving from the outside to the centre:
第一張圖代表沿根狀莖直切的切面圖。生長的端點在右側,在那裡您可以看到單個菌絲,它們相互纏繞但通常都是互相平行的,在這區域,菌絲在功能上沒有差異。往後一點,根狀莖的結構存在顯著差異。從外向中心移動:
The narrow black outer zone represents an outer skin – solid and black in a number of species, but not always
狹窄的黑色外區代表外層表皮——在許多物種中是堅實和黑色的,但並不必然
The dark brown area represents a region packed with narrow hyphae
深啡色區域代表一個充滿幼細菌絲的區域
The speckled blue/light brown area contains many wider hyphae
有斑點的藍色/淺啡色區域包含許多較粗大的菌絲
The second diagram shows a cross-section taken along the red line, well behind the growing apex, in the first diagram. Once again you can see the thin, black protective skin and just in from that the band of narrow hyphae. The area that was a speckled blue/brown in the previous diagram is now shown in more detail, with a mix of narrow and broad hyphae (the hyphal walls shown in blue and the interior white). The broad hyphae are called vessel hyphae and the interiors of these are often empty, allowing the easy flow of nutrients. Within the broad central region the light brown area may contain more narrow hyphae or be composed of glues that bind the hyphae together.
As an example of the variation in rhizomorph structure, the Australian species Armillaria luteobubalina produces rhizomorphs where there is a very large, mostly empty channel constituting a large part of each rhizomorph. It had long been supposed that this channel would allow the efficient transport of oxygen to the advancing apex, but proof of this has come only fairly recently. Laboratory experiments at the University of Sydney and the University of New South Wales have shown that rhizomorphs of Armillaria luteobubalina conduct oxygen. When this species was grown on agar in a petri dish, cylindrical air pores grew vertically above the fungal culture, to about 7mm in height. Rhizomorphs developed below the air pores, grew a short distance down into the agar and then turned to continue growing horizontally. The channels within the air pores were continuous with the central channels in the rhizomorphs and tests showed that oxygen moved through the system. This reference-button takes you to an abstract of a talk about this work, given at the 2002 International Mycological Congress in Oslo. A couple of the people engaged in this work found air-pore-like structures in Norwegian Armillaria-infected wood, during a field excursion associated with the Congress. Those structures, produced by a European species of Armillaria, looked very much like the air pores produced in the laboratory experiments with the Australian Armillaria luteobubalina.
作為根狀莖結構差異的一個例子,澳大利亞的蜜環菌的根狀莖中有一個非常大的、大部分為空心的通道,構成每個根狀莖的大部分。長期以來,人們一直認為這條通道可以有效地將氧氣輸送到生長中的端點,但到了最近這才得以證明。雪梨大學和新南威爾斯大學的實驗室實驗表明了,蜜環菌的根狀莖可以傳導氧氣。當這物種在培養皿中的植物膠上培養時,圓柱形氣孔在真菌培養物上方垂直生長,高度約為7毫米。根狀莖在氣孔下方發育,向下生長進入植物膠一小段距離,然後轉向繼續水平生長。氣孔內的通道與根狀莖的中央通道是連通的,測試表明氧氣在整個系統中流動。這個參考按鈕連結到 2002 年奧斯陸國際真菌學大會上關於這項工作的座談摘要。在與大會相關的實地考察期間,參與這項工作的幾個人在受挪威蜜環菌感染的木材中發現了類似氣孔的結構。這些結構是由歐洲蜜環菌屬產生的,看起來非常像澳大利亞蜜環菌屬實驗室實驗中產生的氣孔。
Armillaria is a world-wide genus of parasitic fungi, with different species in different continents and the species of this genus produce well-structured rhizomorphs. Vessel hyphae have been found in the rhizomorphs of some Armillaria species, but not in Armillaria luteobubalina. The researchers at the universities of Sydney and New South Wales point out that this does not mean that Armillaria luteobubalina rhizomorphs don’t conduct nutrients – just that they don’t have empty vessel hyphae to do the job. In this species, the main role of the rhizomorphs is the transport of oxygen through areas with low oxygen levels – such as moist soils.
蜜環菌是遍及全世界的寄生真菌屬,在不同的大洲有不同的物種,該屬的物種產生結構良好的根狀莖。在一些蜜環菌屬的根狀莖中發現了容器菌絲,但在澳大利亞的蜜環菌中沒有發現。雪梨和新南威爾斯大學的研究人員指出,這並不意味澳大利亞的蜜環菌根狀莖並不傳導養分——只是它們沒有空心的容器菌絲負責這項工作。在這個物種中,根狀莖的主要作用是在低氧含量的區域輸送氧氣——例如在潮濕的土壤中。
Apart from various parasitic fungi, such as Armillaria, many saprotrophic and mycorrhizal species also produce rhizomorphs. The photo shows two immature fruiting bodies of the endemic Australian mycorrhizal species Dermocybe globuliformis, revealed when the surface layer of leaf litter was pulled aside. You can see some diffuse pale yellow mycelium as well as a few, darker yellow rhizomorphs.
除了各種寄生真菌,如蜜環菌,許多腐生菌和菌根菌也會生成根狀莖。照片 顯示了澳大利亞菌根物種禿頭皮菌(Dermocybe globuliformis)的兩個未成熟果體,如將枯枝落葉的表層拉開,就會看得見。你可以看到一些散開的淡黃色菌絲體以及一些深黃色的根狀莖。
Rhizomorphs serve varied purposes. The example of oxygen transport in Armillaria has already been mentioned. Many rhizomorphs can also be looked at as exploratory or migratory structures. For example, suitable food sources for saprotrophic, wood-decay fungi can be widely separated from each other. Look at the fallen trunks or branches in a woodland. Typically there is some distance between them. To you the distances between the different bits of fallen timber may not seem great, perhaps a half-metre here or a few metres there, but to a fungus the intervening areas of bare, wood-free soil may as well be oceans. If such a fungus has exhausted all the nutrients in one piece of wood, it faces a challenge getting to another piece of wood. Rhizomorphs can offer considerable advantages as exploratory tools. The outer skin protects the inner hyphae in the traversal of an inhospitable region. Additionally, rhizomorphs grow much more quickly than isolated hyphae, thereby speeding up the exploration for fresh food sources.
根狀莖有多種功能。之前已提過蜜環菌中運輸氧氣的例子。許多根狀莖也可以視為探索性或遷移性結構。例如,腐生性真菌、腐木真菌的合適食物來源可以廣泛分佈和互相分離。可看看樹林中倒下的樹幹或掉下的樹枝。它們之間通常都有一些距離。對你來說,這些倒下的不同木材之間的距離可能看起來並不大,可能這裡只有半米或那裡有幾米,但對真菌來說,中間空曠的、無木的泥土的區域可算是大海。當這種真菌已經耗盡了一塊木材中的所有養分,那麼它就面臨著需覓得另一塊木材的挑戰。作為探索性工具,根狀莖可以提供相當大的優勢。外層表皮可在穿越荒涼區域時保護裡面的菌絲。還有的是,根狀莖的生長速度比獨立的菌絲快得多,從而加快了對新鮮食物來源的探索。
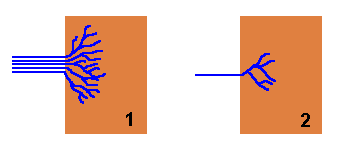
Rhizomorphs also offer considerable advantages when a new piece of wood is found. As there are typically numerous hyphae making up a rhizomorph, when it strikes a new piece of wood the numerous hyphal tips can fan out and rapidly colonise the new wood as a diffuse, branched mycelium. Compare the two figures in the diagram below. In each case the brown rectangles represent pieces of wood. In Figure 1, a rhizomorph has come from the left and found the wood, whereupon the constituent hyphae fan out into a radiating mycelium and fairly quickly have occupied a good proportion of the wood. Figure 2 shows what happens when just a single hypha strikes a similar piece of wood. Once in the wood it will also branch and spread out, but for any time period it will have occupied less of the wood than the rhizomorphic fungus.
當發現一塊新木材時,根狀莖也會提供相當大的優勢。由於根狀莖是由許多菌絲所組成,當它碰到一塊新木材時,許多菌絲導端會散開,並以擴散分支形式的菌絲體迅速殖種在新木材上。 比較以下的兩個圖示。同樣地在兩種情況下,淺啡色的長方形代表木塊。 在圖1中,根狀莖從左邊來到並找到木塊,於是組成莖體的菌絲散開成放射狀的菌絲體,並頗快地佔據了木塊很大的一部分。圖2顯示了當只有一個菌絲發現一塊類似的木塊時會發生什麼事。進入木材後,它也會分支並散開,但無論哪個時段,它所佔據的木材都比根狀莖真菌少。
A well-structured rhizomorph that is able to transport large amounts of water, nutrients or oxygen to the advancing apex will easily fuel the rapid expansion of the fungus into new territory. The contrast between a rhizomorph and an individual hypha is similar to the difference between a multi-lane highway and a narrow, winding road.
結構良好的根狀莖可將大量的水分、養分或氧氣輸送到前進中的根尖,可容易地為真菌提供迅速擴張到新的區域的動力來源。根狀莖和單個菌絲之間的對比就好像多線行車公路和狹窄迂迴的道路之間的分別。
If the new wood is unoccupied by any other fungi or micro-organisms, figure 1 shows that the newly arrived fungus can quickly colonize this pristine territory before it’s found by other organisms. On the other hand, the new piece of wood may already be occupied by other fungi or various micro-organisms, in which case the newly arrived fungus must contend with those existing occupants. If the new fungus has arrived via a rhizomorph then it may be able to overcome the defences of the existing occupants because of the mass of hyphae that are brought to the wood simultaneously. To use a military analogy, figure 1 is equivalent to landing an army division in enemy territory whereas figure 2 is equivalent to landing only a small squad of soldiers in the same hostile territory.
如果新木材沒有被任何其他真菌或微生物佔據,圖1顯示了新到達的真菌可以在其他生物發現它之前,迅速在這片嶄新領土上殖種。另一方面,這新的木材可能已經被其他真菌或各種微生物佔據,在這種情況下,新來的真菌必須與那些既有的佔據者競爭。如果新真菌是以根狀莖的形式抵達,那麼它可能能夠克服既有的佔據者的防禦,因為大量的菌絲同時抵達木材。使用一個軍事類比,圖1就等於在敵方領土上登陸一個師的軍隊,而圖2就等於在同一敵方領土上只登陸一小隊士兵。
Armillaria luteobubalina is one example of rhizomorphs enabling a fungus to traverse inhospitable areas. A further example of this is the Dry Rot fungus (Serpula lacrymans ), a species that can cause significant damage to timber in damp, poorly ventilated buildings. The rhizomorphs produced by this fungus can extend for several metres across brickwork in the search for fresh wood and the ability to form such rhizomorphs makes this a very destructive fungus. As long as there is a source of water and nutrients, the rhizomorphs can transport these over considerable distances and across inhospitable terrain to support the advancing mycelium. There is, once again, a very apt military analogy. As long as there are both a well-stocked supply depot and a secure, large-capacity supply line, a considerable front-line force can be supported over a long distance.
澳大利亞的蜜環菌是根狀莖使真菌能夠穿越荒涼區域的的例子。另一個例子是木腐真菌 (Serpula lacrymans ),這種真菌會在潮濕、通風不良的建築物中對木材造成嚴重損壞。在尋找新鮮木材時,這種真菌產生的根狀莖可在磚牆上延伸數米,這種根狀莖的能力使其成為一種極具破壞性的真菌。只要有水和養分的來源,根狀莖就可以將它們運送到相當長的距離並穿越荒涼的地帶來支持菌絲體的前進。再一次,有一個非常恰當的軍事類比。只要有充足的補給庫和安全的大容量補給線,就可以遠距離支援相當大的前線部隊。
In all of the above examples, the rhizomorphs have been more-or-less at ground level, but they can also be found off the ground. In tropical rainforests a considerable amount of the leaf and twig litter that falls from the taller trees is often trapped, at least temporarily, in understorey trees and shrubs. Numerous wiry rhizomorphs (to 1mm in diameter) wind their way through the trapped litter. A study in Ecuador found 200 to 300 kg (dry weight) of such trapped litter per hectare, containing at least 500 to 600 metres of rhizomorph per kilogram of litter. Another way of reporting that rhizomorph abundance is to say that there’s at least 100 kilometres of rhizomorph per hectare. A number of common rhizomorph-producing fungi were in the genera Marasmius or Marasmiellus. Many of the species in these genera produce tiny mushrooms as fruiting bodies, with the caps often under a centimetre in diameter. The rhizomorphs hold on to fallen leaves and twigs by producing splayed-out adhesion pads. Once these pads have a secure hold on the plant debris, hyphae can penetrate and fan out as a more diffuse mycelium to extract the nutrients inside. You can find out a bit more about this study in the reference given in the button. Incidentally, the author also reported that the rhizomorphs are so common that th ey are used in Hummingbird nests – which can later produce crops of tiny mushroom.
在以上所有的例子中,根狀莖或多或少都是生在地面上,但在地面以上都可以找到。在熱帶雨林中,從較高的樹木落下的大量樹葉和樹枝枯枝經常被困在下層植被和灌木中,即使只是暫時性的。無數細長的根狀莖(直徑可達1毫米)蜿蜒穿過被困的枯落枝葉。厄瓜多爾的一項研究發現,每公頃有200至300公斤(乾重)的此類被困的枝葉,而每公斤枝葉至少帶有500至600米的根狀莖。另一種簡述根狀莖密度的方法是說每公頃至少有100公里的根狀莖。許多常見的可產生根狀莖的真菌屬於小皮傘(Marasmius)或微皮傘(Marasmiellus)屬。這些屬中的許多物種產生細小的蘑菇作為果體,菌蓋的直徑通常不到一厘米。根狀莖通過產生張開的粘附墊來粘住落葉和樹枝。當這些墊子牢固地固定在植物殘骸之上,菌絲就可以滲透並散開,成為更擴散的菌絲體提取內裡的養分。您可以在這按鈕中連結到的參考資料中找到有關這項研究的更多信息。順 帶一提,作者還報告中提到,這種根狀莖非常普遍地被用於蜂鳥巢——而之後可以生產出大量的小蘑菇。
Fairy rings
精靈環
As already noted, a mycelium growing in the ground does not expand as a disk but as a ring, with most of the activity near the outer edge of the ring. Not surprisingly, mushrooms often appear to grow in a ring, the so-called fairy ring that reflects the underground mycelial presence. The most commonly seen rings are probably those produced by Marasmius oreades (which even gets the common name of Fairy Ring Mushroom). The mushrooms produced by this fungus do not have a striking appearance, generally being pale brown and often only 2 or 3 centimetres in diameter (though sometimes up to 5 centimetres). However, the mushrooms often appear in large numbers, in obvious rings, in open grassy areas such as sports fields and lawns in homes and public parks. These pictures show both the mushrooms in close view and views of a couple of fairy rings.
如前所述,在地下生長的菌絲體不會像圓盤一樣擴張,而是環狀擴張,大部分活動都在環的外圍附近。不出所料,蘑菇通常呈環狀生長,即反映地下菌絲體存在的所謂精靈環。最常見的環大多是由硬柄小皮傘(Marasmius oreades)所生產的環(它甚至得到了精靈環蘑菇的俗名)。這種真菌生產的蘑菇沒有很特別的外觀,一般呈淺啡色,直徑通常只有2或3厘米(但有些可達5厘米)。然而,這種蘑菇經常大量出現,呈明顯的環狀,出現在開闊的草地上,如家庭和公園的草坪和運動場。這些照片既顯示了蘑菇的近距離樣貌,也顯示了幾個精靈環的樣貌。
Marasmius oreades is a widespread species, found in numerous countries in both hemispheres, and may appear at almost any time of the year (even summer) after some rain. While this may be the most commonly seen ring-forming species, several species of Agaricus (the genus that includes the Field Mushrooms) as well as a few puffball and puffball relatives are also fairly often seen in rings. While the rings produced by these fungi are often easily noticed because they often occur in open, grassy areas, many other fungi are known to produce their fruiting bodies in rings. However, it’s easy to miss the ring arrangement in a bushy forest area.
硬柄小皮傘(Marasmius oreades)是一種遍佈各地的物種,在兩半球的許多國家都有發現,並且可能在一年中的幾乎任何時間(甚至夏季)在下過雨後都會出現。雖然這可能是最常見的環狀生長物種,但也經常可在環中找到幾種傘菌屬(包括野蘑菇在內的屬)以及一些擔子菌和擔子菌近親的蘑菇。雖然這些真菌產生的環通常很容易被注意到,因為它們經常出現在開闊的草地區域,但許多其他真菌亦會產生環狀的果體。然而,在茂密的森林區域中它們的環狀的排列很容易被錯過。
The rate of growth of fairy rings varies, even within the one species. For example, several European studies of Marasmius oreades fairy ring growth showed a variety of average annual radial growth rates, from ten centimetres per year to thirty-five centimetres per year.
精靈環的生長速度不盡相同,即使在同一物種內也如是。例如,歐洲對硬柄小皮傘(Marasmius oreades)精靈環生長的幾項研究表明,它們每年平均徑向增長率各不相同,從每年10厘米到每年35厘米不等。
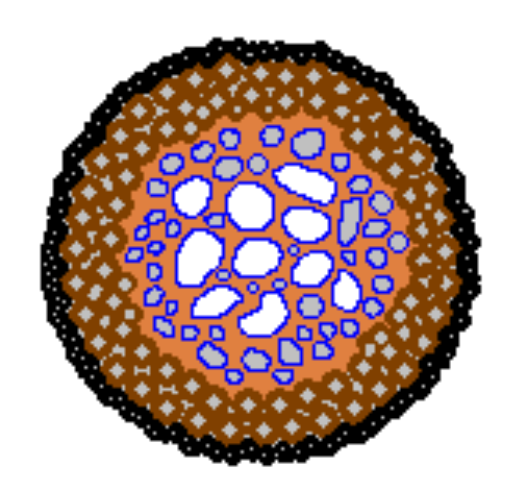
In some cases the ring-like underground mycelium has little or no visible effect and only makes its presence known when it produces the visible fruiting bodies. On the other hand, Marasmius oreades is an example of a fungus where the mycelium has a visible effect throughout the year, often showing up as a ring of brownish or dead grass, with two zones of greener, more vigorous grass immediately inside and outside the ring of dead grass. The following diagram shows a stylized aerial view of such a ring.
在某些情況下,環狀地下菌絲體幾乎或完全沒有可見的蹤跡,只有當它生產出可見的果體時才會知道它的存在。另一方面,硬柄小皮傘(Marasmius oreades)是一種其菌絲體的存在全年都可明顯被看見的真菌的一個例子,通常可見的狀態是一圈啡色或已枯萎的死草環,緊貼其內圍和外圍有兩個更綠、更有生氣的草區。下圖顯示了這種環的模擬鳥瞰圖。
What is the reason behind these zones of differing grass growth? The advancing edge of the underground mycelium secretes chemicals that break down organic matter in the soil to release nutrients for the fungus to use. For a short time the grass just beyond the outer edge of the mycelium can also make use of these nutrients, in particular nitrogen – which makes the grass much greener than the grass further ahead of the outer edge of the mycelium. A little way behind the outer edge, the mycelium is much denser, actively feeding on the liberated nutrients and leaving little for the grass. Moreover, the mycelium may be impervious to water, thereby reducing the water available to the grass and so putting further stress on the grass. Further inwards, there are the remnants of the cannibalized mycelium. This is broken down by various micro-organisms and the resulting nutrients are released into the soil. This increase in nutrients behind the mycelial ring explains the enhanced grass growth immediately behind the ring of brownish grass. Therefore the brownish grass recovers as the ring expands. The second diagram shows a vertical view, into the soil, along the dotted blue line in the first diagram.
這些區域中的草的不同生長狀態的背後原因是什麼?地下菌絲體生長的前沿會分泌化學物質分解土壤中的有機物質,釋放出可供真菌使用的養分。在一段短時間內,菌絲體外圍以外的草也可以利用這些養分,尤其是氮——這使得這裡的草比起離菌絲外圍更遠的草更綠。在外圍後面一點的位置,菌絲體密度要大得多,積極地吸收已釋放的養分,留下給草可吸收的會很少。此外,菌絲體可能並不透水,從而減少了草可用的水,而對草增加了更多的壓力。再往裡面,殘餘了被拆解的菌絲體。這會被各種微生物分解,而所產生的養分會被釋放到土壤中。菌絲環後面養分的這增加解釋了緊接在啡色草環後面的草的生長有所增強。因此,隨著環的擴大,啡色的草會恢復。第二張圖顯示了沿著第一張圖中的藍色虛線切入土壤的垂直切面圖。
Many of the fungi that form fairy rings are saprotrophic species but mycorrhizal fungi can also form fairy rings. In the case of a mycorrhizal fungus, the mycelium is always attached to the roots of one (or more) plants. Such a fairy ring is called a tethered ring whereas the rings formed by the unattached (non-mycorrhizal) mycelia, such as those of Marasmius oreades, are called free rings. The photo shows a tethered ring formed by a species of Cortinarius in a Eucalyptus forest at Tidbinbilla Nature Reserve, in the Australian Capital Territory.
許多形成精靈環的真菌是腐生性物種,但菌根真菌也可以形成精靈環。菌根真菌的話,菌絲體總是附著在一種(或多種)植物的根上。這種精靈環稱為繫繩環,而由獨立(非菌根)菌絲體形成的環,例如硬柄小皮傘(Marasmius oreades)的那些,稱為游離環。這照片 顯示了在澳大利亞首都區的提賓比拉(Tidbinbilla)自然保護區的桉樹林中由一種絲膜菌(Cortinarius)形成的繫繩環。
Sclerotia
菌核
A sclerotium is a mass of compacted hyphae, commonly more or less spherical in shape and often with a dark, tough outer skin. Sclerotia are produced by many fungi and range in size from under a millimetre to over 20 centimetres in diameter. A sclerotium is a resting structure that allows a fungus to sit out the hard times, analogous to (but clearly different from) a plant bulb or corm. The outer skin protects the internal hyphal mass from drying out. When conditions improve, the sclerotium may produce a new mycelium or fruiting bodies. Two examples are Hypholoma tuberosum and Laccocephalum mylittae , formerly called Polyporus mylittae. The sclerotium of Laccocephalum mylittae has been used as a food by the Australian aborigines. Species in the genus Morchella are further examples of sclerotium-producing fungi.
菌核是一團壓縮緊實的菌絲,通常大概呈球形,外皮通常都是深色且堅韌的。 菌核由許多真菌產生,直徑從一毫米以下到超過20厘米不等。菌核是一種休止結構,可以讓真菌度過艱難時期,類似於(但明顯不同於)植物的鱗莖或球莖。外皮保護內部菌絲團免於乾涸。當環境改善時,菌核可能會產生新的菌絲體或果體。其中有兩個例子是瘤核垂幕菇(Hypholoma tuberosum) 和雷丸(Laccocephalum mylittae) ,以前稱為多孔菌(Polyporus mylittae)。雷丸(Laccocephalum mylittae)的菌核以前被澳大利亞原住民用作食物。羊肚菌屬(Morchella) 中的物種是產生菌核的真菌的另一個例子。
This drawing shows a tropical polypore Lignosus rhinocerus. The dotted line represents ground level and you can see that the fruiting body has grown out from a buried sclerotium.
此圖顯示了熱帶的多孔虎乳芝(Lignous rhinocerus)。虛線代表地面,可以看到果體從藏在地下的菌核中生長出來。
Sclerotia have been described as similar to the noduli, the primordial swellings that give rise to the fruiting bodies. Indeed one mycologist has written that sclerotia could be described as arrested and sometimes greatly oversized noduli. A pseudosclerotium is a heterogeneous mix of hyphae and substrate material. Sometimes the distinction can be difficult:
菌核被描述為類似於小結核(Noduli),是生長出果體的原初腫塊。的確如此,一名黴菌學曾把菌核描述為被抑制,有時會是超大的小結核。偽菌核是菌絲和基底物料的異質混合物。有時候並不容易區別:
Pseudosclerotia begin their development by delimitating a usually rather large volume of substrate, wood or soil, by surrounding it with dense hyphal strands or sheaths. In most cases, hyphae grow out of the sheath into the delimitated chunk of substrate and slowly transform the latter into an intimate mixture of fungal biomass and residual substrate. When this happens in wood, such as felled trees or living roots, the wood inside the pseudosclerotium is protected against decay by other microorganisms and serves as a food reserve. In Wolfiporia extensa a sclerotium first grows around a root, then penetrates into it, and the resulting intermediate stage is a pseudosclerotium. Then the wood is completely digested and replaced by fungal mass. When this happens, the resulting body is a sclerotium. This means that the proportion of substrate in a pseudosclerotium varies enormously, from almost zero to more than half the body. Therefore the limit between sclerotia and pseudosclerotia is often blurred.
偽菌核會通過認定和以緊密的菌絲綑索和以外層覆套把通常相當大量的基底、木或泥土來開始發展。在大多數情況下,菌絲從覆套中向外生長到認定的基底堆,再緩慢地把它們轉化為真菌生長所需的生物物質群和殘餘基底的緊密混合物。當這種情況發生在木材上,例如被砍伐的樹木或有生命的根部,偽菌核內的木材會受到其他微生物的保護而不會腐爛,而成為食品儲備。在茯苓(Wolfiporia extensa)的例子,菌核會先在樹根周圍生長,然後再滲透進入它的內部,而這過渡性階段出現的就是偽菌核。然後將木材會被完全消化並成為真菌質群。當這種情況發生時,形成的質體就是菌核。這意味著在偽菌核中的基底所佔的比例可以有很大差異,從近乎零到超過整體的一半以上。因此,菌核和偽菌核之間的界線通常都很模糊。
The pseudosclerotium of Polyporus tuberaster, a northern hemisphere species, was long a puzzle, for here was something, apparently a stone, that produced fungi. It was noted in medieval times by Mathaeus Silvaticus, who referred to it as lapis lincis (literally ‘lynx stone’) in his pharmacopeia, Pandectarum medicinae. He wrote that it was created from the coagulation of lynx urine and noted that it could be used to produce fungi throughout the year. Be aware that various Classical authors refer to lynx stones but the evidence points to these being references to amber, not to sclerotia or pseudosclerotia. After Mathaeus’ time the pseudosclerotium was known by various names, incorporating at least one of ‘lynx’ or ‘stone’ and three examples are Lapis lyncurius, Fungus lyncurius and Pietra fungaja. The first to publimonographsh a good study of this pseudosclerotium was Pier Antonio Micheli, in his 1729 Nova plantarum genera, and this PDF presents both his illustrations and his words. The first modern examination of this fungus was published in 1845 by Carl Brunner and this PDF contains one of Brunner’s plates showing several polypores growing from a pseudosclerotium.
北半球物種菌核多孔菌(Polyporus tuberaster)的假菌核長期以來都是一個謎,因為那裡有些東西,顯然是石頭,可以生產出真菌。馬菲雅斯‧薜弗迪卡士(Mathaeus Silvaticus)在中世紀已對它作了記載,他在他的藥典《醫藥總覽》(Pandectarum medicinae)中將其稱為lapis lincis(字面意思是「猞猁石」)。他寫道,它是由猞猁的尿液凝固而成,並指出它可以全年都可生長出真菌。請注意,許多古典學者都提到了猞猁石,但有證據表明這些是指琥珀,而不是菌核或偽菌核。在馬菲雅斯的時代之後,偽菌核曾被稱為不同的名稱,當中都包含了「猞猁」或「石頭」一詞,其中三個例子是寶石(Lapis lyncurius)、寶菌(Fungus lyncurius)和菌石(Pietra fungaja)。皮雅‧安東尼奧‧米切尼(Pier Antonio Micheli) 在他 1729 年的專論《新植物屬》(Nova plantarum genera) 中第一個發表了對這些偽菌核的很好的研究,這PDF 展示了他的插圖和文字。卡爾·布倫納(Carl Brunner)則於1845 年發表了對這種真菌的第一次現代研究,這PDF 包含了布倫納 (Brunner) 的一個圖板,當中顯示了從假菌核中生長出的幾個多孔菌。
Dendropolyporus umbellatus, another northern hemisphere polypore, produces dense clusters of fruiting bodies from pseudosclerotia which can be coralloid or very contorted in shape, as shown in this PDF .
豬苓(Dendropolyporus umbellatus)是另一種北半球的多孔菌,會從偽菌核生長出密集的果體叢,其形狀可以是珊瑚狀或非常曲彎的,就如這PDF 中所顯示。
 Collybia tuberosa is a sclerotium-producing mushroom genus, also found in the northern hemisphere. The mushrooms develop from the decayed remains of other fungi and a hand-coloured, 1820s illustration (as well as some additional information) is included in this PDF .
球莖金錢菌(Collybia tuberosa)是一種會生產菌核的蘑菇屬,亦是在北半球被發現。 這蘑菇從其他真菌的腐化殘骸中發育而成,這PDF 中包含了1820年代的手繪插圖(以及一些附加信息)。
Lentinus tuberregium, shown in this PDF , is found in many tropical areas, including the Australian tropics, and produces sclerotia.
這PDF 中顯示的球莖香菇(Lentinus tuberregium)在於許多熱帶地區有發現,包括澳大利亞熱帶地區,亦會生產菌核。
Longevity
長壽
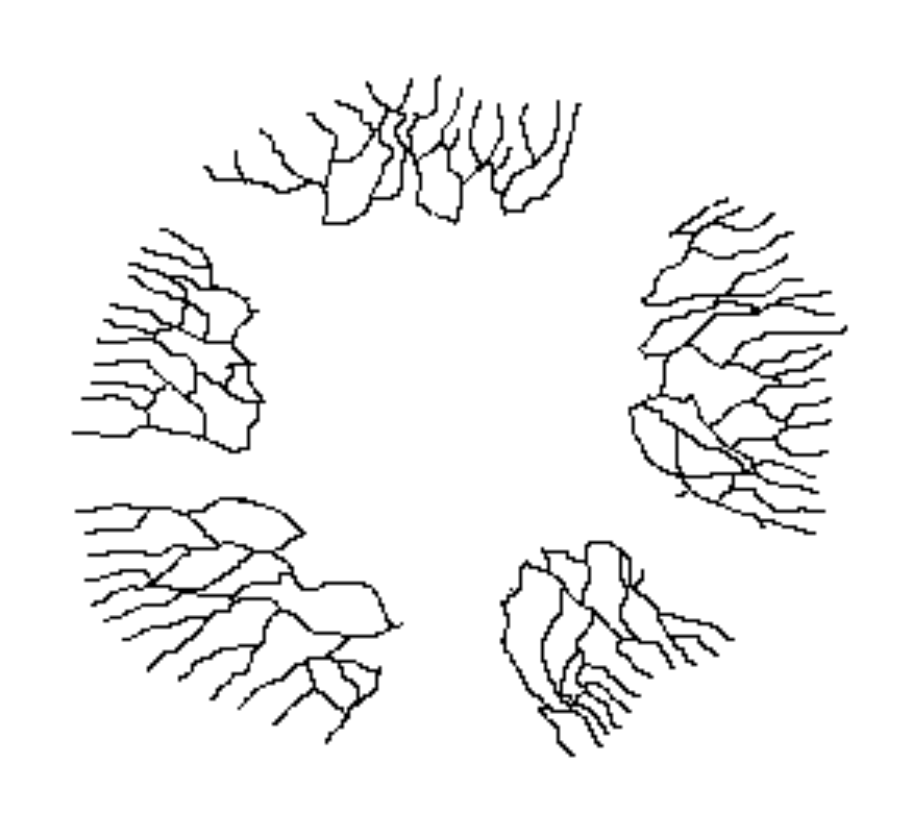
In principle, a mycelium that can spread through the ground could keep on growing indefinitely and large, old mycelia are known. Of course, once a mycelium has spread to a considerable size it becomes more likely that it will suffer disruptions and fragmentation. For example, a landslip, local flooding or antagonistic organisms could destroy sections of the mycelium, leaving isolated fragments where once there was a continuous mycelium.
原則上,可以在地上擴展的菌絲體可以無止境地繼續生長和擴大,亦有發現古老的菌絲體。當然,一旦菌絲體擴散到相當大的規模,它就更有可能遭受破壞和碎片化。 例如,山泥傾瀉、局部性洪水或敵對性生物都可能會破壞部分的菌絲體,在曾經一體連結的菌絲體的地方留下孤立的碎片。
This is illustrated by the diagram, which is simply a copy of an earlier diagram, with several parts now erased to mimic disturbances of one kind or another. The five parts of the original mycelium can continue growing independently but, though no longer physically connected, they are still genetically identical and can be looked at as the one individual. As they continue to grow it is possible that at some future stage two of these currently disconnected mycelia will come into contact. Then, since genetically identical mycelia can fuse, the two mycelia can re-join.
這幅圖說明了這種情況,它是上面一幅圖的抄本,這裡擦除了幾個部分以模擬不同形式干擾。原本菌絲體分裂出的五個部分可以繼續獨立生長,但雖然它們不再有物理上的直接聯繫,但它們在基因上仍然是一樣的,可以視為一個個體。隨著它們繼續生長,在將來的某個時候,這些當前斷開的菌絲體中的其中兩個可能會發展出接觸。然後,由於基因相同的菌絲體可以融合,兩個菌絲體可以重新結合。
Genetic testing answers the question of whether two separated mycelia are the one individual and in this way various large, old mycelia have been found. An example of large size and great age is provided by an Armillaria ostoyae mycelium in Oregon. Armillaria produces mushrooms as fruiting bodies and the genus is present in all continents, though with different species in different areas. The species of Armillaria are plant parasites and many species spread from tree to tree via rhizomorphs that grow through the soil. The particular Armillaria ostoyae mycelium in question is spread over 890 hectares and is estimated to be at least 2,400 years old. While this individual may hold the size/age record, there are other documented instances of fungal mycelia with impressive sizes and ages – when compared to the human scale. Mycelia spread over several tens of hectares, and with estimated ages of several hundred to a thousand years, are not rare.
基因測試回答了兩個分離的菌絲體是否是一個個體的問題,並通過這種方式發現了不同的巨大和古老的菌絲體。 俄勒岡州的一個奧氏蜜環菌菌絲體提供了一個大面積和年齡久長的例子。蜜環菌會生產蘑菇果體,這個屬在各大洲都有,儘管在不同地區有不同的物種。蜜環菌屬植物寄生物種,許多通過在泥土中生長的根狀莖從一棵樹擴展到另一棵樹。以上提及的那個蜜環菌菌絲體擴展達 890公頃以上,估計至少有2,400年的歷史。雖然這個個體可能擁有面積/年齡的記錄,但與人類標準相比,還有其他有記載的真菌菌絲體的大小和年齡都令人印象深刻。擴展連達數十公頃,估計年齡在幾百到一千年之間的菌絲體,並不罕見。
On the other hand, consider species such as Cordyceps gunnii and Poronia erici . The first is a parasite of larvae of Ghost Moths in the genus Oxycanus and the second is a specialist dung fungus, found in the dung of various herbivores. A couple of things in common to these two species is that their substrates are very limited in size and separated in space. For example, while there are many Oxycanus individuals in Australian woodlands or forests, they are not in constant physical contact with one another. Similarly, there are numerous piles of herbivore droppings in Australian grasslands, woodlands and forests – but they are separated from one another. Once the Cordyceps mycelium has spread throughout its host and extracted all the nutrients – it has nowhere else to go. Similarly, the Poronia mycelium has nowhere to go once it has spread through the dung. These species do not produce hyphae or rhizomorphs that will spread through the soil looking for other larvae or dung piles. Once the nutrients are exhausted these species produce fruiting bodies and rely on spores. In the case of Cordyceps, any spores that get into fresh larvae will start a new cycle of infection. Poronia spores that land on nearby grass blades will be eaten by herbivores and later pass out in their droppings.
另一方面,就岡恩蟲草(Cordyceps gunnii) 和多孔石南(Poronia erici) 等物種的情況。第一個是奧斯簡洛斯(Oxycanus)屬的鬼蛾幼蟲的寄生菌,第二個是一種特殊的糞便真菌,可在草食動物的糞便中發現。這兩個物種的一些共同點是它們的基底尺寸非常有限並且互相分隔。例如,雖然澳大利亞林地或森林中有許多奧斯簡洛斯(Oxycanus)個體,但它們之間並沒有持續的直接接觸。同樣,在澳大利亞的草原、林地和森林中也有無數的的草食動物糞便堆——但它們彼此分隔。一旦蟲草菌絲體已遍布整個宿主並提取了所有養分後——它就無處可去。同樣,多孔石南菌絲體一旦已遍布糞便堆後就無處可去。這些物種不會生長出可在泥土中擴展以尋找其他幼蟲或糞堆的菌絲或根狀莖。當養分已耗盡,這些物種就會生長出果體並依賴孢子傳播。就蟲草而言,任何可進入新鮮幼蟲的孢子就會開始新的感染週期。落在附近草葉上的多孔石南孢子會被草食動物吃掉,然後在它們的糞便中排出。
In conclusion
總結
The English mycologist Alan Rayner has spent many years studying the behaviour of mycelia in forests. In the introduction to the reference given in the button, he wrote:
英國真菌學家艾倫·雷納(Alan Rayner)花了多年時間研究菌絲體在森林中的行為。在按鈕連結到的參考文獻的簡介中,他寫道:
“…there is perhaps a natural tendency to regard …the mycelium as a boring, uniformly absorbent mass of hyphal threads which, whilst important in energy capture, only really become interesting when parts of it aggregate and differentiate into a fruit body…… I have increasingly come to regard the mycelium as a heterogenous army of hyphal troops, variously equipped for different roles and in varying degrees of communication with one another. Without a commander, other than the dictates of their environmental circumstances, these troops organise themselves into a beautifully open-ended or indeterminate dynamic structure that can continually respond to changing demands. Recall that during its potentially indefinite life, a mycelial army may migrate between energy depots; absorb easily assimilable resources such as sugars; digest refractory resources such as lignocellulose; mate, compete and do battle with neighbours; adjust to changing microclimatic conditions; and reproduce.”
「……可能有一種都可算是自然的傾向,認為……菌絲體是一種無聊的、均勻吸收的菌絲線束,雖然它在能量吸收方面很重要,但只有當它的一部分聚合並分支成果體時才真正變得有趣 …… 我越來越認為菌絲體是一支由菌絲組成的異質軍隊,裝備各異,用於不同的角色,彼此之間的交流程度也有所不同。沒有指揮官,除了它們身處的環境條件的規限,這些部隊將自己組織成一個美麗的開放式或不確定的動態結構,可以不斷回應不斷變化的需求。回想一下,在其可能是無限的壽命期間,菌絲體軍隊可能會在能量庫之間遷移;吸收好像糖那些容易同化的資源、消化好像木質纖維素那些難處理的資源、與近鄰交配,競爭和戰鬥、適應不斷變化的微氣候條件;並繁殖。」
Wonderful blog post. This is absolute magic from you! I have never seen a more wonderful post than this one. You’ve really made my day today with this. I hope you keep this up!

breaks down parts of the hyphal walls in the mycelium and uses the released nutrients in fruit body production.

presents both his illustrations and his words. The first modern examination of this fungus was published in 1845 by Carl Brunner and this PDF




Thanks for such a great post and the review, I am totally impressed! Keep stuff like this coming.
It’s going to be finish of mine day, but before end I am reading
this impressive post to improve my experience.
What a really awesome post this is. Truly, one of the best posts I’ve ever witnessed to see in my whole life. Wow, just keep it up.
Always so interesting to visit your site.What a great info, thank you for sharing. this will help me so much in my learning
I have read your excellent post. This is a great job. I have enjoyed reading your post first time. I want to say thanks for this post. Thank you…
Wonderful blog post. This is absolute magic from you! I have never seen a more wonderful post than this one. You’ve really made my day today with this. I hope you keep this up!
Thank you for your post, I look for such article along time, today i find it finally. this post give me lots of advise it is very useful for me.